DNA TECHNOLOGY AND GENOMICS Part I
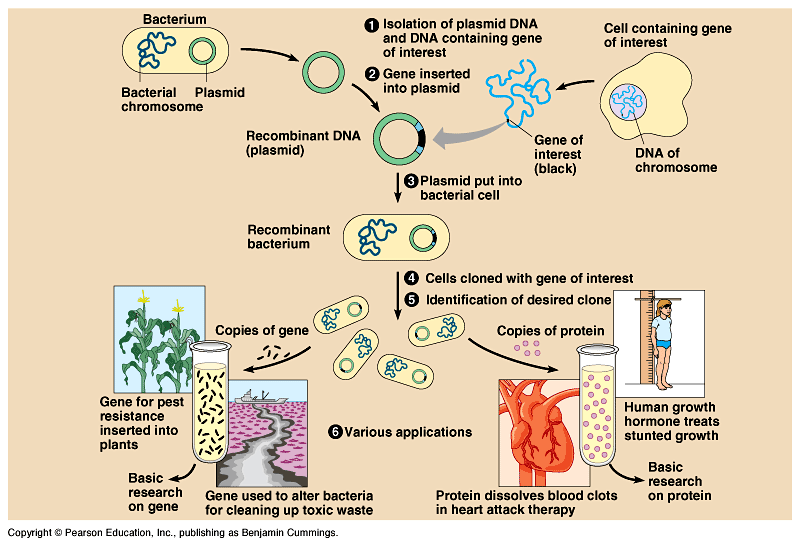
DNA Cloning - Fig
20.1
To study a particular gene, you need to isolate only the small,
well-defined, portion of a chromosome containing the gene.
Gene cloning makes multiple identical copies of gene-sized
pieces of DNA.
- A foreign gene is inserted into a bacterial plasmid and this
recombinant DNA molecule is returned to a bacterial cell.
Every time this cell reproduces, the recombinant plasmid is replicated
as well and passed on to its descendents.
Under suitable conditions, the bacterial clone will make the
protein encoded by the foreign gene.
Uses of cloned genes:
To produce a protein product, like human
growth hormone.
To prepare many copies of the gene itself.
to determine
the gene's nucleotide sequence
- to provide
an organism with a new metabolic capability by transferring a
gene from another organism.
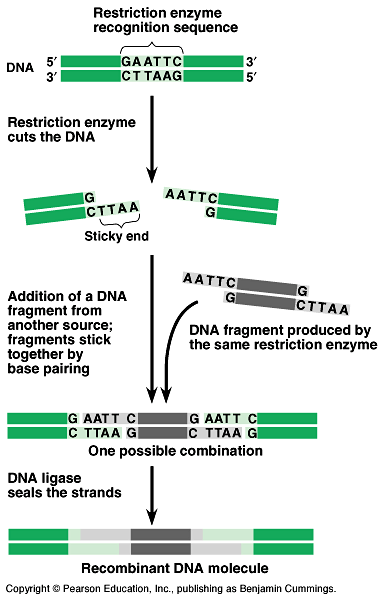
Restriction enzymes are used to make recombinant DNA - Fig
20.2.
- Restriction enzymes cut DNA molecules at specific
locations called restriction sites.
In nature, bacteria use restriction enzymes to cut foreign DNA
and protect their own DNA by methylation.
Restrictions enzymes recognize short DNA nucleotide sequences
and cut at specific point in these sequences.
Because the target sequence usually occurs many times on a long
DNA molecule, an enzyme will make many cuts.
Copies of a DNA molecule will always yield the same set of restriction
fragments when exposed to a specific enzyme.
Restriction enzymes leave sticky ends.
These will bond with complementary single-stranded stretches
on other DNA molecules cut with the same restriction enzyme.
DNA ligase seals the strand.
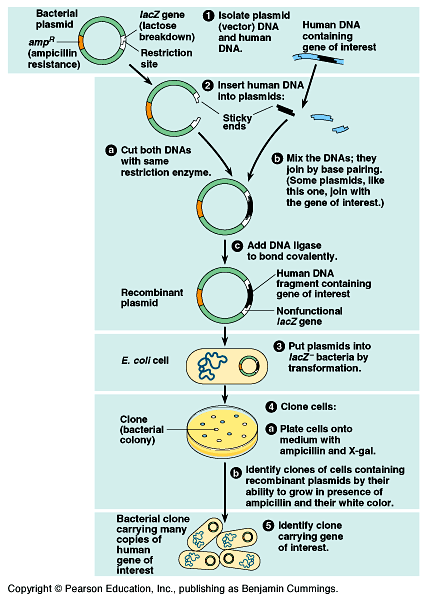
Genes can be cloned in recombinant DNA vectors - Fig
20.3.
- Recombinant plasmids are produced by splicing restriction
fragments from foreign DNA into plasmids.
The original plasmid is called a cloning vector.
The process of cloning a human gene in a bacterial plasmid
can be divided into five steps.
1) Isolation of vector and gene-source
DNA.
- The source DNA comes from human tissue cells.
The source of the plasmid is typically E. coli.
This plasmid carries two useful genes, ampR, conferring
resistance to the antibiotic ampicillin and lacZ, encoding
the enzyme beta-galactosidase which catalyzes the hydrolysis
of sugar.
2) Insertion of DNA into the vector.
- same restriction enzyme
- sticky ends
DNA ligase.·
3) Introduction of the cloning vector
into cells.
- Bacterial cells that are are lacZ- take up the recombinant
plasmids by transformation.
Some bacteria have taken up the desired recombinant plasmid DNA,
other bacteria that have taken up other DNA.
4) Cloning of cells (and foreign genes).
- Plate out the transformed bacteria on a solid nutrient medium
containing ampicillin and a sugar called X-gal.
Only bacteria that have the ampicillin-resistance plasmid will
grow.
- The X-gal in the medium is used to identify plasmids that
carry foreign DNA.
Bacteria with plasmids lacking foreign DNA stain blue when beta-galactosidase
hydrolyzes X-gal.
Bacteria with plasmids containing foreign DNA are white because
they lack beta-galactosidase.
5) Identifying cell clones with the right
gene.
- Sort through the bacterial colonies with foreign DNA to find
those containing the gene of interest.
Nucleic acid hybridization, depends on base-pairing between
the gene and a complementary sequence, a nucleic acid probe,
on another nucleic acid molecule. Fig
20.4.
A radioactive or fluorescent tag labels the probe.
The probe will hydrogen-bond specifically to complementary single
strands of the desired gene.
After denaturation (separating) the DNA strands in the
plasmid, the probe will bind with its complementary sequence,
tagging colonies with the targeted gene.
Inducing a cloned eukaryotic gene to function in a prokaryotic
host can be difficult so employ an expression vector, a
cloning vector containing the requisite prokaryotic promotor
upstream of the restriction site.
The presence of introns creates problems for expressing
these genes in bacteria. Fig.
20.5.
- To express eukaryotic genes in bacteria, a fully processed
mRNA acts as the template for the synthesis of a complementary
strand using reverse transcriptase.
This complementary DNA (cDNA), with a promoter,
can be attached to a vector for replication, transcription, and
translation inside bacteria.
Can use eukaryotic cells as host for cloning and expressing
eukaryotic genes
- Yeast cells, single-celled fungi, are as easy to grow
as bacteria and have plasmids, rare for eukaryotes.
Scientists have constructed yeast artificial chromosomes
(YACs) - an origin site for replication, a centromere,
and two telomeres -with foreign DNA.
These chromosomes behave normally in mitosis and can carry more
DNA than a plasmid.
Eukaryotic hosts are capable of providing the posttranslational
modifications that many proteins require.
Several techniques facilitate entry of foreign DNA.
- electroporation
microscopically thin needles.
For plants, DNA is attached to microscopic metal particles and
fired into cells with a gun.
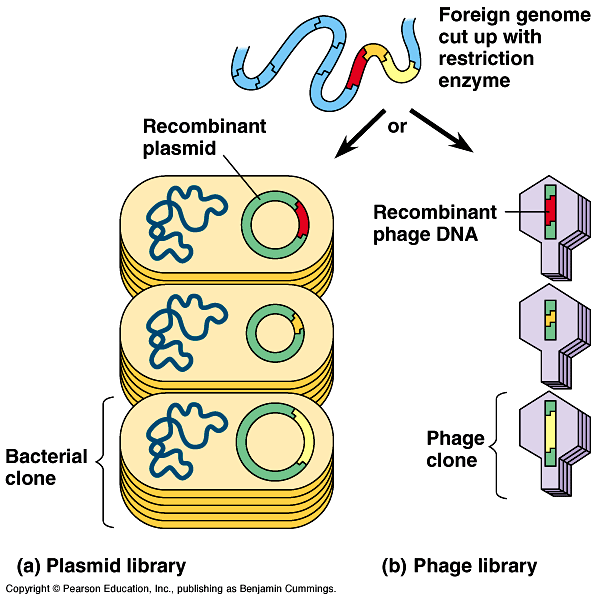
Cloned genes are stored in DNA libraries - Fig
20.6
- A complete set of recombinant plasmid clones, each carrying
copies of a particular segment from the initial genome, forms
a genomic library.
The library can be saved and used as a source of other genes
or for gene mapping.
Certain bacteriophages are also common cloning vectors
for making libraries.
Fragments of foreign DNA can be spliced
into a phage genome using a restriction enzyme and DNA ligase.
The recombinant phage DNA is packaged
in a capsid in vitro and allowed to infect a bacterial
cell.
Infected bacteria produce new phage particles,
each with the foreign DNA.
The polymerase chain reaction (PCR) clones DNA entirely
in vitro - Fig.
20.7
- DNA cloning is the best method for preparing large quantities
of a particular gene or other DNA sequence.
When the source of DNA is scanty or impure, the polymerase
chain reaction (PCR) is quicker and more selective.
The DNA is incubated in a test tube with special DNA polymerase,
isolated from bacteria living in hot springs, a supply of
nucleotides, and short pieces of single-stranded DNA
as a primer.
PCR can make billions of copies of a targeted DNA segment in
a few hours, much faster than cloning with recombinant bacteria.
In PCR, a three-step cycle-heating, cooling, and replication-brings
about a chain reaction that produces an exponentially growing
population of DNA molecules.
Examples:
Fragments of ancient DNA from a 40,000-year-old
frozen woolly mammoth.
DNA from tiny amount of blood or semen found
at the scenes of violent crimes.
DNA from single embryonic cells for rapid prenatal
diagnosis of genetic disorders.
DNA of viral genes from cells infected with
difficult-to-detect viruses such as HIV.
DNA Analysis and Genomics
- Comparisons among whole sets of genes and their interactions
is the field of genomics.
One indirect method of rapidly analyzing and comparing genomes
is gel electrophoresis. Fig
20.8.
Gel electrophoresis separates macromolecules - nucleic
acids or proteins - on the basis of their rate of movement through
a gel in an electrical field.
Rate of movement depends on size, electrical charge, and other
physical properties of the macromolecules.
For linear DNA molecules, separation depends mainly on size,
with longer fragments migrating less along the gel.
Restriction fragment analysis detects DNA differences that
affect restriction sites - Fig
20.9.
- After treating long DNA molecules with a restriction enzyme,
the fragments can be separated by size via gel electrophoresis.
This produces a series of bands that are characteristic of the
starting molecule and that restriction enzyme.
The separated fragments can be recovered undamaged from gels,
providing pure samples of individual fragments.
We can use restriction fragment analysis to compare two different
DNA molecules representing, for example, different alleles, which
may differ in one or more restriction sites.
If they do differ in restriction sites, each will produce different-sized
fragments when digested by the same restriction enzyme.
- The restriction fragments from the two alleles will produce
different band patterns.
Restriction fragment analysis is sensitive enough to distinguish
between two alleles of a gene that differ by only base pair in
a restriction site.
Although electrophoresis will yield too many bands to distinguish
individually, we can use nucleic acid hybridization with
a specific probe to label discrete bands. The radioactive label
on the single-stranded probe can be detected by autoradiography.
To compare 3 individuals:
- Add the restriction enzyme to each of the three samples to
produce restriction fragments.
Separate the fragments by gel electrophoresis.
Southern blotting (Southern hybridization) allows us to
transfer the DNA fragments from the gel to a sheet of nitrocellulose
paper, still separated by size. Fig 20.10.
This also denatures the DNA fragments.
Bathing this sheet in a solution containing the probe allows
the probe to attach by base-pairing (hybridize) to the DNA sequence
of interest and then visualize bands containing the label with
autoradiography.
For our three individuals, the results of these steps show that
individual III has a different restriction pattern than individuals
I or II.
Southern blotting can be used to examine differences in noncoding
DNA as well.
Differences in DNA sequence on homologous chromosomes that
produce different restriction fragment patterns are called restriction
fragment length polymorphisms (RFLPs) and can serve
as a genetic marker for a particular location (locus) in the genome.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}